 Reviews
Reviews
Trilobites: Ancient Arthropods of the Paleozoic Seas
Trilobites are among the most iconic and scientifically important fossils ever discovered. These extinct marine arthropods lived for an astonishing 270 million years, thriving from the early Cambrian Period (about 521 million years ago) until their final disappearance at the end of the Permian Period (252 million years ago). For comparison, dinosaurs existed for roughly 165 million years—meaning trilobites endured for far longer than the age of dinosaurs.
Over 20,000 described species of trilobites are known, and new species continue to be discovered. They ranged in size from tiny forms only a few millimeters long to giants exceeding 70 centimeters (over 2 feet), such as Isotelus rex, the largest trilobite ever found. Trilobites occupied nearly every marine ecological niche imaginable, from shallow coastal waters to deep-sea environments.

Trilobites: Ancient Arthropods of the Paleozoic Seas
Their fossils are so abundant and diverse that trilobites are often called the “index fossils of the Paleozoic.” Geologists use them to date and correlate rock layers across continents. Despite their long success and complexity, trilobites left no direct living descendants, making them a fully extinct branch of the arthropod tree. Beyond their scientific value, trilobites are prized by collectors for their intricate exoskeletons, dramatic spines, compound eyes, and exceptional preservation.
What makes trilobites especially fascinating is how much they reveal about early animal life: the evolution of complex eyes, the development of segmented bodies, and the rise of advanced behaviors hundreds of millions of years before vertebrates dominated the seas.
Scientists recognize 8 or 9 major trilobite orders, depending on classification scheme. Commonly recognized orders include Redlichiida, Ptychopariida, Asaphida, Phacopida, Lichida, Odontopleurida, Corynexochida, Harpetida, and Proetida, reflecting the extraordinary evolutionary diversity of the group.
Trilobites are extinct members of the phylum Arthropoda, the same broad group that includes insects, spiders, crabs, and lobsters. They belong to their own class, Trilobita, and are not direct ancestors of any living animal, though they share many traits with modern arthropods. The name trilobite comes from the Greek tri (three) and lobos (lobe), referring to the three longitudinal sections of their bodies.
Trilobite anatomy is both highly standardized and remarkably adaptable, reflecting their long evolutionary history and ecological diversity. Their hardened exoskeleton was composed primarily of calcite and chitin, providing protection, structural support, and a surface for muscle attachment.
Trilobite bodies are divided into three main regions:
Cephalon (Head)
The cephalon, or head, functioned as the control center of the trilobite. It typically housed a pair of compound eyes when present, a single pair of antennae used for sensory perception, and the mouth along with its associated feeding appendages. The central raised area of the cephalon is known as the glabella, which often overlies the stomach and digestive organs. Variations in the shape of the glabella are important for identifying trilobite species and may also reflect differences in feeding strategy.
Thorax (Articulated Body Segments)
The thorax consisted of multiple articulated segments, ranging from just a few to more than twenty depending on the species. These segments allowed flexibility for walking, burrowing, and enrolling. Each thoracic segment bore a pair of limbs beneath it.
Pygidium (Tail)
The pygidium was formed from fused segments and varied greatly in size. In some trilobites it was tiny (micropygous), while in others it was large or even larger than the head (macropygous). Its size and shape are key classification features.
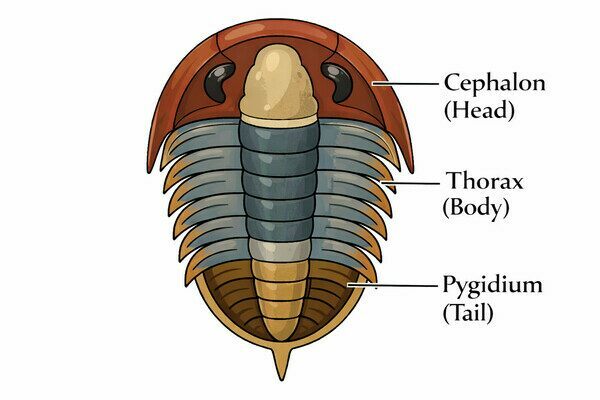
Trilobite Body Plan: Cephalon, Thorax and Pygidium
Viewed from above, trilobites are divided lengthwise into:
A central axial lobe, which housed vital organs
Two pleural lobes on either side, which aided in movement and protection
This tri-lobed arrangement is one of the defining features of the group.
For much of scientific history, trilobite limbs were unknown, but exceptional fossil preservation has revealed that trilobites possessed biramous (two-branched) appendages, similar to those of some modern crustaceans.
Each body segment typically bore a pair of limbs consisting of:
An inner walking leg used for locomotion and handling food
An outer branch, likely functioning as a gill for respiration
These limbs suggest that trilobites were capable walkers on the seafloor and, in some cases, weak swimmers. Limb structure was surprisingly conservative across trilobite groups, even as exoskeleton shapes diversified.
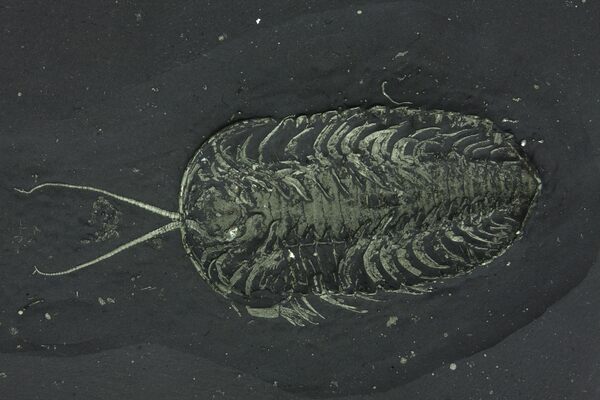
A ventrally preserved, Triarthrus trilobite from New York with rare soft-bodied preservation showing legs, gills and antennae.
Trilobites had a single pair of antennae attached beneath the cephalon. These flexible, segmented structures were likely used to sense the environment, locate food, and navigate the seafloor. Antennae are rarely preserved but are known from sites such as the Burgess Shale and Chengjiang Biota.
Trilobite eyes are among the earliest complex eyes known in the fossil record. Most species possessed compound eyes made of calcite lenses—a unique feature rarely seen in modern animals. A few modern exceptions include brittlestars (ophiuroids), which use calcite lenses in their skeletal plates to sense light, and certain deep-sea crustaceans, which incorporate calcite into eye structures for light guidance rather than true image formation.
There were several eye types:
Holochroal eyes – many small lenses under a single cornea (most common)
Schizochroal eyes – fewer, larger lenses, each with its own cornea (found in phacopid trilobites)
Abathochroal eyes – rare and structurally unique
Some trilobites were completely blind, suggesting life in deep water or low-light environments.

A schizochroal eye of the trilobite Eldredgeops rana crassituberculata of Ohio.
One of the most visually striking aspects of trilobite morphology is the presence of spines, tubercles, ridges, and other forms of ornamentation. These features vary enormously between groups and became increasingly elaborate over time.
Early Cambrian trilobites were generally smooth-bodied or only lightly ornamented. Prominent spines became more common during the Ordovician and especially widespread in the Silurian and Devonian, coinciding with the rise of increasingly effective predators such as jawed fishes and large cephalopods.
Possible Functions of Spines
Trilobite spines likely served multiple purposes, including:
Defense against predators: Long pleural and axial spines made trilobites more difficult to swallow or manipulate.
Stabilization on soft substrates: Spines may have helped distribute body weight, preventing sinking into soft sediment.
Camouflage and disruption of outline: Irregular shapes could make trilobites less recognizable to predators.
Species recognition and display: In some cases, ornamentation may have played a role in mating or species differentiation.

A super spiny Comura bultyncki trilobite from Morocco covered in long spines.
Trilobites Known for Spines and Ornamentation
Some trilobite groups are especially famous for their elaborate armor:
Lichida – heavily built trilobites with thick exoskeletons and prominent spines
Odontopleurida – extreme spinosity, including long, delicate spines covering much of the body
Phacopida – generally robust trilobites, some with modest spines and strong enrollment
Certain Cambrian redlichiids – early experiments in genal and thoracic spines
In contrast, smooth-bodied trilobites such as many asaphids and proetids relied more on enrollment, size, or ecological positioning for protection rather than extreme ornamentation.
The evolution of spines and ornamentation in trilobites provides a clear example of an evolutionary "arms race" between predators and prey during the Paleozoic, recorded in stone across hundreds of millions of years.
The original coloration of trilobites is not preserved in most fossils, as pigments break down during fossilization. However, paleontologists can make informed inferences about trilobite color based on comparisons with modern arthropods, ecological context, and rare exceptional preservation.
Most trilobites were likely colored in subdued tones such as browns, greens, grays, or mottled patterns that would have provided camouflage against the seafloor. Species living in shallow, well-lit environments may have exhibited more complex patterning to break up their outline, while deep-water or burrowing trilobites were likely uniformly dark or pale.
Some researchers have suggested that certain trilobites—particularly spiny or visually prominent forms—may have displayed contrasting patterns or highlights, either for species recognition or deterrence. However, bright warning coloration is considered unlikely for most species, given their benthic lifestyles and reliance on camouflage.
Rare fossil sites preserve subtle evidence of original surface texture, and in a few cases, differences in mineral replacement hint at possible pigment distribution, though these interpretations remain speculative.
Overall, trilobites were probably not brightly colored in the way many modern reef animals are, but instead displayed practical, environment-specific coloration well suited to survival in Paleozoic seas.
Hypostome – A hard plate located on the underside of the cephalon that covered the mouth. The shape and attachment of the hypostome are important indicators of feeding behavior, with some types suggesting scavenging and others active predation.
Glabella – The raised central region of the cephalon, often overlying the stomach and digestive organs. Its shape varies widely among trilobites and is a critical identification feature.
Genal spines – Spines extending from the rear corners of the cephalon. These may have aided in defense, stabilization on soft sediment, or enrollment.
Pleural lobes – The lateral sections of the trilobite body on either side of the axial lobe.
Pleural spines – Extensions from the pleural lobes, especially common in spiny trilobites, likely serving defensive and stabilizing functions.
Axial lobe – The central longitudinal lobe of the trilobite body, containing many vital organs.
Thoracic segments – The articulated body segments between the cephalon and pygidium that allowed flexibility and enrollment.
Pygidium – The tail shield formed from fused posterior segments; its size and shape are key taxonomic features.
Enrollment – The defensive behavior of curling the body into a ball to protect soft tissues.
Like all arthropods, trilobites possessed a rigid, mineralized exoskeleton that could not grow continuously. To increase in size, they underwent a process known as molting (ecdysis), in which the old exoskeleton was periodically shed and replaced with a larger one. This occurred many times throughout a trilobite’s life, from tiny larval stages to full adulthood.
Molting in trilobites typically involved the opening or separation of specific parts of the exoskeleton—most commonly along facial sutures on the head or at the junction between the head (cephalon) and thorax. The trilobite would wriggle free of the old shell, leaving behind an empty exoskeleton that often retained its original shape. After emerging, the animal’s new exoskeleton was initially soft and vulnerable, hardening over time as it mineralized.
Many trilobite fossils actually represent molted exoskeletons rather than dead individuals, which helps explain their remarkable abundance in the fossil record. These molts are often recognizable by disarticulated parts, missing cheeks, or repeated occurrences of similarly sized specimens in the same layers.
Trilobites continued molting until reaching their final growth stage, after which they underwent a terminal or final molt. This last molt marked the end of growth and the onset of full maturity. In some species, the final molt coincided with changes in body proportions or the development of specialized features, such as exaggerated spines or ornamentation, possibly related to reproduction or defense. Once the final molt was complete, the trilobite would no longer shed its exoskeleton for the rest of its life.
Paleontologists distinguish trilobite molts from actual carcasses by examining how the fossil is preserved. Molted exoskeletons often show signs of deliberate separation, such as missing free cheeks, an opened head shield, or a gap between the head and thorax—common exit points used during molting. These remains may also appear flattened, slightly disarticulated, or repeated in large numbers at a single site, reflecting normal growth rather than mass death.
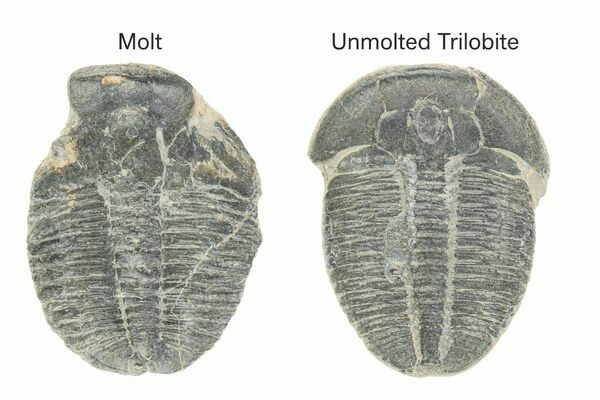
Two Elrathia trilobite fossils, the one on the left being a molt while the one on the right is the actual dead individual.
In contrast, trilobite carcasses are more likely to be fully articulated, with the head, thorax, and tail preserved in life position. Some show evidence of enrollment (curling up), a defensive posture unlikely during molting. The surrounding sediment and orientation can also provide clues, helping scientists determine whether a specimen represents a shed exoskeleton or the preserved remains of the animal itself.
Trilobites occupied a remarkable range of ecological roles on the ancient seafloor, with their modes of life closely intertwined with how they obtained food. Although the majority of trilobites were benthic—living on or just beneath the seafloor—their lifestyles were far from uniform, reflecting millions of years of adaptation to different marine environments.
Many trilobites lived as epifaunal crawlers, spending their lives moving across the surface of the sediment. These forms, common among groups such as ptychopariids and early redlichiids, likely foraged constantly as they crept over the seafloor. They fed primarily as scavengers and detritivores, consuming organic debris, decaying organisms, and microbial mats that blanketed Cambrian and Ordovician seabeds. Their anatomy was well suited to this lifestyle: moderately flexible bodies allowed them to navigate uneven substrates, while their limbs could manipulate sediment and food particles beneath them.
Other trilobites adopted a more concealed existence as shallow infaunal burrowers. These species were adapted to pushing through soft sediments, where they may have searched for buried organic material or small invertebrates. Smooth exoskeletons, reduced spines, and streamlined outlines helped minimize resistance as they moved beneath the surface. Living partially buried would have offered protection from predators while granting access to nutrient-rich sediments, making detritivory and opportunistic scavenging particularly effective feeding strategies in this niche.
A smaller but significant number of trilobites appear to have ventured above the seafloor. Some asaphids and other forms may have been capable of short bursts of swimming or hovering just above the substrate. Flattened bodies and well-developed pleural lobes would have aided this semi-nektonic lifestyle. These trilobites may have combined benthic feeding with the ability to relocate quickly or exploit suspended food resources, such as fine organic particles in the water column.

Cyclopyge is a rare, free-swimming trilobite distinguished by its enormous, downward-facing eyes. Its sleek, streamlined body indicates an active life in the open water column instead of along the ocean floor.
While scavenging and detritivory were the most widespread feeding strategies, trilobites were not limited to passive feeding. Certain robust and heavily ornamented species—particularly among the lichids—likely acted as active predators. With strong cephalic structures, spines for grasping or defense, and powerful appendages, these trilobites may have hunted small worms, soft-bodied arthropods, or other vulnerable invertebrates on or within the sediment.
Some trilobites may also have practiced filter feeding, using specialized appendages to strain suspended particles from the water, especially in calm, shallow marine environments. This strategy would have complemented a more stationary or slow-moving lifestyle, allowing trilobites to exploit nutrient-rich waters without constant locomotion.
Underlying these ecological roles was a strong link between form and function. The shape of the glabella and cephalon is thought to reflect feeding behavior, with broader or more expanded glabellas often associated with more active or specialized feeding strategies. Together, these anatomical and behavioral adaptations reveal trilobites not as uniform, primitive arthropods, but as highly diverse and ecologically sophisticated animals that played many roles in Paleozoic marine ecosystems.
Enrollment—curling the body into a tight ball—was one of the most important defensive behaviors in trilobites. This behavior protected vulnerable soft tissues and concealed limbs beneath the exoskeleton. Trilobites with well-developed articulation and reinforced margins, such as phacopids, were especially adept at enrolling.
Many species also evolved long spines, thick armor, or ornamented exoskeletons, which served to deter predators or make handling more difficult.

A perfectly enrolled Morocops trilobite.
Trilobites were a major food source in Paleozoic marine ecosystems and faced predation throughout their history. Likely predators included:
Anomalocaridids: Large Cambrian arthropods, such as Anomalocaris, are thought to have preyed on early trilobites, as evidenced by bite marks and damaged exoskeletons.
Cephalopods: By the Ordovician and later periods, straight-shelled and coiled cephalopods likely hunted trilobites.
Jawed fishes: During the Silurian and Devonian, the rise of armored and jawed fishes dramatically increased predation pressure on trilobites.
Other arthropods: Larger arthropods may also have preyed upon smaller trilobite species.
Fossil evidence such as repaired injuries, bite marks, and abnormal growth patterns provides direct insight into predator–prey interactions.

A Utaspis trilboite from Utah with a large bite mark. Possibly left by an Anomalocaridid.
Some fossil deposits contain mass mortality layers of trilobites, suggesting group behavior such as seasonal migration, synchronized molting, or breeding aggregations. Trackways preserved in sediment show multiple individuals moving in the same direction, supporting the idea of coordinated movement.
Such behaviors may have helped trilobites avoid predators, find mates, or exploit seasonal food resources.

A line of Lonchodomas trilobites showing potential migrating behavior.
Direct evidence of trilobite reproduction is rare, but several important discoveries—particularly from Silurian and Ordovician deposits in New York State—provide valuable insight. Fossils from New York have yielded clusters of small, rounded structures preserved in close association with trilobite specimens, most notably with enrolled individuals of genera such as Flexicalymene. These clusters are widely interpreted as eggs, based on their size, shape, repeated occurrence, and spatial relationship to the adult trilobites.
In addition, exceptionally preserved specimens from New York have been interpreted to show paired internal structures consistent with reproductive organs, suggesting that some trilobites may have carried or brooded eggs rather than releasing them freely into the water column. This supports the idea that at least some trilobites practiced parental investment, a behavior seen in certain modern arthropods.
Together, these fossils indicate that trilobite reproduction was likely more complex than once assumed, involving egg production, possible brooding, and seasonal reproductive behavior tied to migration or molting cycles.
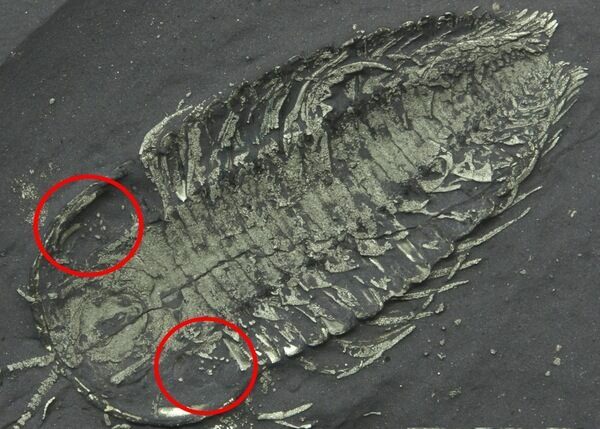
A triarthrus trilobite with what has been interpretted as eggs. Interestingly these are in the same location of the body where horseshoe crabs carry their eggs.
Trilobites possess one of the richest and most informative fossil records of any extinct animal group. Their mineralized exoskeletons fossilized readily, and because they were abundant, widespread, and rapidly evolving, trilobites are invaluable for understanding Paleozoic geology and life. Trilobite fossils are found on every continent, including Antarctica, reflecting the global distribution of ancient seas.
The way trilobites are preserved depends largely on sediment type, burial conditions, and post-depositional geology.
Flattened trilobites are typically found in fine-grained shales and mudstones. Compaction during burial compresses the exoskeleton, producing fossils preserved in two dimensions. Many classic Cambrian trilobites from North America and China are preserved this way.
Three-dimensionally preserved trilobites are usually found in limestones or dolomites, where early mineralization prevented compaction. These specimens often retain natural curvature, relief, and fine detail, making them especially prized by collectors.
Exceptional preservation can also occur when trilobites are rapidly buried by sediment flows or preserved in low-oxygen environments that inhibit decay.
Trilobite fossils have a truly global footprint, with discoveries spanning every continent and representing hundreds of millions of years of Earth’s history. From ancient seabeds preserved in fine detail to fossil-rich layers that capture extraordinary diversity, certain trilobite localities stand out above the rest. In this article, we highlight some of the world’s most important trilobite sites, chosen for the exceptional preservation of their specimens, the remarkable variety of species they yield, or their long-standing significance to fossil collectors and researchers alike.
Morocco is widely regarded as the global epicenter of modern trilobite collecting and preparation, earning its reputation as the mecca for trilobite enthusiasts. The Anti-Atlas Mountains are especially famous for producing extraordinarily ornate Devonian trilobites, including large phacopids, lichids, and spiny odontopleurids that rank among the most visually striking trilobites ever discovered. In addition to these Devonian faunas, Morocco also hosts extensive Ordovician and Cambrian deposits, yielding large asaphids, early trilobite forms, and a remarkable continuity of trilobite history across much of the Paleozoic.
What makes Morocco truly unique is the combination of geology and human tradition. Vast regions of the Anti-Atlas expose fossil-bearing strata over enormous distances, with minimal vegetation, erosion cover, or tectonic faulting, allowing trilobite-rich layers to be traced and worked for miles. Thousands of people are involved in the commercial excavation and preparation of trilobites, often using highly refined techniques to extract fossils from hard limestone and reveal exquisite detail. As a result, Morocco is the source of the majority of trilobite specimens available on the global fossil market today, making it unmatched in both productivity and influence within the trilobite collecting community.
The Burgess Shale occupies a foundational place in the history of paleontology and trilobite research. Discovered in 1909 by Charles D. Walcott, this Cambrian fossil deposit—dated to approximately 508 million years ago—was among the first sites to demonstrate that delicate soft tissues could be preserved alongside mineralized skeletons. At the time of its discovery, this challenged long-held assumptions about the limits of fossil preservation and dramatically expanded scientific understanding of early animal life. Specimens preserving appendages, digestive systems, and respiratory structures provided the first compelling evidence that trilobites were complex, fully articulated animals rather than simple, rigid shells. These fossils laid the groundwork for modern interpretations of trilobite locomotion, feeding behavior, and ecological roles during the Cambrian explosion.
Today, the Burgess Shale is strictly protected, and collecting is prohibited, with material limited to museum collections and historically excavated specimens.
The Great Basin region of Utah and Nevada is one of the most important trilobite areas in the world, with hundreds—if not thousands—of Cambrian species described, reflecting an extraordinary diversity during the early evolution of trilobites. Well-known Middle Cambrian deposits such as the Wheeler Shale and Marjum Formation in Utah’s House Range, along with Lower Cambrian strata near Pioche, Nevada, have been studied for over a century and continue to be actively collected by hundreds of amateur and professional fossil hunters each year.
Oklahoma holds a special place in trilobite collecting history, best known for its exquisitely preserved Devonian trilobites from the Haragan Formation, which have been collected and studied for decades. Localities such as Black Cat Mountain have produced spectacularly spiny odontopleurids and amazing phacopids, alongside a diverse assemblage of other Devonian forms, while lesser-known Ordovician and Silurian deposits reveal a surprisingly rich trilobite fauna spanning much of the Paleozoic. Oklahoma is also significant for its role in the evolution of fossil preparation, as Bob Carroll helped develop and popularize modern trilobite preparation techniques there—methods that have since become standard practice and elevated trilobite collecting into both a scientific and artistic discipline.

Collecting trilobites from the Haragan Formation in Oklahoma.
The Ordovician trilobite localities near St. Petersburg, Russia are among the most important classical trilobite sites in the world and have played a major role in the early development of trilobite research and taxonomy. Dating to roughly 485–444 million years ago, these deposits—often preserved in limestone and shale—have yielded abundant and well-studied trilobites, particularly large asaphid forms such as Asaphus, Megistaspis, and related genera. Many of these trilobites are notable for their broad, smooth bodies and, in some species, dramatically elevated eyes adapted for low-light environments.
These Russian Ordovician localities were heavily collected and described during the 19th and early 20th centuries, forming the basis for much of the foundational literature on Ordovician trilobites. Their exceptional abundance and preservation have made them critical for biostratigraphy and evolutionary studies, especially in understanding early trilobite gigantism and morphological specialization.
New York State is one of the most historically important regions in the world for trilobite research, with classic fossil localities spanning the Ordovician and Silurian periods. The state is home to the famous Beecher’s Trilobite Beds (Upper Ordovician, ~445 million years old), a globally significant Lagerstätte where trilobites such as Triarthrus eatoni are preserved with soft tissues, including legs and antennae, through pyritization. These specimens provided some of the earliest direct evidence of trilobite limb anatomy and were critical to understanding trilobites as living arthropods rather than simple mineralized shells. New York is also renowned for the Walcott–Rust Quarry, famous for producing beautifully preserved 3D trilobites like Isotelus, Ceraurus and Flexicalymene
Ontario is one of the most important regions in North America for Ordovician trilobites, preserving rich marine faunas dating to roughly 470–450 million years ago. Limestone formations across southern Ontario have yielded abundant trilobites such as Isotelus, Flexicalymene, Ceraurus, and Asaphus, including Isotelus rex, one of the largest trilobite species ever discovered and the official provincial fossil of Ontario. These deposits record shallow, warm Ordovician seas and have played a significant role in understanding trilobite diversity, morphology, and early Paleozoic marine ecosystems.
Historically, Ontario was a productive region for both research and collecting, with many classic museum specimens originating from active quarries. However, collecting has become increasingly difficult in recent decades due to quarry closures, restricted access, and stricter regulations. As a result, new material is far less common today, and Ontario trilobites are now valued primarily for their scientific and historical importance rather than abundance.
Trilobites have fascinated scientists, collectors, and the public for more than two centuries, playing a central role in the development of paleontology itself. Early researchers laid the foundation for our understanding of these ancient arthropods, none more influential than Charles Doolittle Walcott, whose discovery of the Burgess Shale in 1909 revealed exquisitely preserved Cambrian trilobites and other soft-bodied organisms. Before his famous discovery of the Burgess Shale in 1909, Walcott conducted some of his earliest and most important trilobite research at the Walcott–Rust Quarry near Trenton Falls, New York. Walcott’s work dramatically expanded scientific views of early animal life and helped establish trilobites as key indicators of early evolutionary complexity.

Modern day collection of trilobites from the hard limestones of the Walcott-Rust Quarry in New York
Throughout the 19th and 20th centuries, trilobites became a focus for pioneering paleontologists studying evolution, extinction, and ancient marine ecosystems. Their rapid evolutionary turnover and wide geographic distribution made them ideal subjects for biostratigraphy, allowing scientists to correlate rock layers across continents and reconstruct Earth’s deep-time history with remarkable precision.
In the modern era, the field has been equally shaped by the contributions of private collectors, preparators, and dedicated amateurs, particularly in regions such as Morocco, Utah, New York, and the American Midwest. Advances in preparation techniques—especially fine mechanical air abrasion—have revealed delicate features like compound eyes, spines, enrollment mechanisms, and even traces of soft anatomy. Many of the world’s finest trilobite specimens, including fully articulated and three-dimensionally preserved examples, come from the efforts of skilled private preparators working closely with researchers and museums.
This collaboration between collectors, commercial fossil preparators, academic institutions, and museums has led to the discovery of numerous new species and a deeper understanding of trilobite anatomy, behavior, and ecology. Today, trilobites remain indispensable to scientific research, helping illuminate patterns of evolution, mass extinction, and environmental change across hundreds of millions of years.
Beyond their scientific value, trilobites hold immense cultural and aesthetic significance. Their extraordinary diversity—ranging from smooth, streamlined forms to wildly spined and ornamented species—has made them one of the most sought-after fossils in the world. Equal parts scientific tool, historical artifact, and natural work of art, trilobites continue to inspire curiosity and admiration, securing their place as one of the most iconic fossils ever discovered.

An amazingly prepared Erbenochile trilobite from Morocco showing that these extrodinary fossil arthropods can also be natural works of art.
Trilobites first appeared abruptly in the early Cambrian Period as part of the Cambrian Explosion, a time of rapid evolutionary experimentation when most major animal groups first emerged. Although no single direct ancestor has been definitively identified, trilobites are generally thought to have evolved from earlier arthropod-like stem groups.
Potential ancestors include soft-bodied Cambrian arthropods such as Spriggina, Parvancorina, and other members of the broader arthropod stem lineage, though these relationships remain debated. Fossils from sites like the Chengjiang Biota show transitional arthropods with segmented bodies, paired appendages, and developing cephalization—key traits that would later be fully expressed in trilobites.

A fossil of Spriggina floundensi, a possible ancestor of trilobites that lived during the Late Ediacaran, approximately 555 million years ago.
The sudden appearance of trilobites with complex eyes and mineralized exoskeletons suggests that their lineage had already undergone significant evolutionary development before entering the fossil record.
During the Cambrian, trilobites rapidly diversified into numerous forms and ecological roles. Early orders such as Redlichiida and Ptychopariida dominated Cambrian seas. These trilobites exhibited a wide range of body sizes, segment counts, and spinosity, reflecting intense evolutionary experimentation. Cambrian trilobites were primarily benthic crawlers and scavengers, but even at this early stage they occupied multiple niches. Frequent molting and rapid reproduction likely contributed to their success.
The Ordovician Period marked a second major diversification event for trilobites. Many Cambrian groups declined or disappeared, while more derived orders such as Asaphida rose to prominence. Trilobites became larger on average and expanded into deeper and more offshore environments. This period also saw increasing specialization, including eye reduction in deep-water forms and enhanced visual systems in shallow-water species. Trilobites were major components of increasingly complex marine ecosystems during the Ordovician.
The end-Ordovician mass extinction, driven largely by global cooling and sea-level fluctuations, caused significant trilobite losses. Many families and genera vanished, particularly those adapted to shallow, warm-water environments. However, several lineages survived by occupying deeper or more flexible ecological niches.
Following the Ordovician extinction, trilobites rebounded modestly during the Silurian. Orders such as Phacopida and Lichida diversified, often evolving robust bodies, improved enrollment mechanisms, and elaborate spines.
The Devonian Period brought increased predation pressure from rapidly evolving jawed fishes. In response, many trilobites evolved heavier armor, tighter enrollment, and defensive ornamentation. Despite these adaptations, overall trilobite diversity declined throughout the Devonian. The Late Devonian extinction events eliminated many remaining trilobite groups, leaving only a few resilient lineages.

During the Devonian trilobites increasingly evolved long spines, likely a defensive mechanism against predators like jawed fish.
By the Carboniferous Period, trilobites were reduced to a handful of families within the order Proetida. These trilobites were generally small, conservative in form, and often inhabited deeper or more restricted marine environments. Proetids survived multiple environmental fluctuations, including changes in sea level and oxygen availability, demonstrating remarkable resilience despite low diversity.
The final extinction of trilobites occurred during the Permian–Triassic mass extinction, approximately 252 million years ago—the most severe extinction event in Earth’s history. Massive volcanic eruptions, ocean acidification, widespread anoxia, and extreme climate change devastated marine ecosystems. All remaining trilobites, including the last proetids, were wiped out during this event. Unlike earlier extinctions, there was no recovery or rebound for the group.
Although trilobites left no direct descendants, their evolutionary legacy is profound. They provide critical insight into the early evolution of arthropods, the development of complex sensory systems, and the long-term dynamics of extinction and survival. Their rise and fall illustrate both the power and fragility of evolutionary success over geological time.
Trilobites are traditionally divided into a set of major orders based on overall body plan, eye structure, thoracic segmentation, and evolutionary relationships. Depending on the classification scheme used, scientists recognize eight primary trilobite orders, with some debate over additional or historically used groupings.
The order Redlichiida includes some of the earliest and most primitive trilobites known from the fossil record. Redlichiids first appeared in the Early Cambrian (approximately 521 million years ago) and persisted into the Middle Cambrian, disappearing by around 509 million years ago as more derived trilobite groups rose to prominence. They were an important component of early Cambrian marine ecosystems during the initial radiation of trilobites.
Redlichiid trilobites are typically characterized by large, crescent-shaped cephalons, numerous thoracic segments, and relatively small pygidia, giving them an elongated, flexible body plan. Many species possess long, sweeping genal spines extending from the rear of the cephalon, features that may have provided stability on soft substrates or offered some protection from early predators.
These trilobites are generally interpreted as active benthic crawlers, moving across the seafloor in search of food. Their morphology suggests a mobile lifestyle, and they are thought to have been among the earliest trilobites to explore a variety of shallow marine environments. Redlichiids represent some of the earliest evolutionary experiments in trilobite body plans, laying the groundwork for later, more specialized forms.

A Redlichiid trilobite (Olenellus nevadensis) from the Lower Cambrian of Nevada.
The order Ptychopariida represents one of the most diverse, abundant, and long-ranging groups of trilobites in the fossil record. Ptychopariids first appeared in the Early Cambrian (approximately 521 million years ago) and persisted into the Late Ordovician, with some lineages extending into the earliest Silurian. The group reached its greatest diversity and ecological importance during the Cambrian Period, when trilobites dominated marine ecosystems worldwide.
Members of this order typically possess moderate-sized cephalons, well-developed thoraxes composed of numerous segments, and a generally simple, conservative body plan. While often lacking the extreme spinosity or ornamentation seen in other trilobite orders, this morphological consistency proved highly successful and allowed ptychopariids to exploit a wide range of environments.
Ptychopariid trilobites occupied diverse ecological niches, ranging from shallow continental shelf settings to deeper offshore environments. Most are interpreted as benthic detritivores or scavengers, living on or within soft seafloor sediments. Their adaptability and wide geographic distribution made them one of the most successful trilobite groups of the Paleozoic.
Because of their abundance, rapid evolutionary turnover, and broad distribution, ptychopariids are extremely important for biostratigraphy, particularly in Cambrian-aged rocks where they are widely used to define and correlate stratigraphic zones.

The Ptychopariid trilobite (Elrathia kingii) from the Wheeler Shale in Utah is probably the most recognizable trilobite in the world due to millions of them having been collected.
The order Corynexochida includes a diverse assemblage of trilobites distinguished by their forward-tapering (conical) glabellas, a feature that often gives the cephalon a pointed or wedge-shaped appearance. Many corynexochids also bear prominent thoracic or cephalic spines, though the degree of ornamentation varies widely across the group.
Corynexochid trilobites first appeared during the Early Cambrian (approximately 521 million years ago) and persisted into the Ordovician Period, with some lineages extending into the early Silurian. The group reached its greatest diversity during the Cambrian, when trilobites were undergoing rapid evolutionary radiation, and remained an important component of marine faunas into the Ordovician before declining.
Morphologically, Corynexochida encompasses both smooth-bodied, streamlined forms and heavily ornamented species with long, sweeping spines, reflecting a wide range of ecological strategies. These trilobites are generally interpreted as active benthic animals, living on or near the seafloor. Some species show features consistent with predatory or scavenging behavior, while others may have been semi-infaunal, partially burrowing into soft sediments for feeding or protection.

A Corynexochid trilboite (Olenoides nevadensis) from the Middle Cambrian deposits of Utah.
The order Asaphida includes a diverse group of often large, smooth-bodied trilobites that were especially successful during the Ordovician Period. Asaphids are typically characterized by a wide, flattened body, a broad pygidium, and a reduced number of thoracic segments, giving many species a streamlined, almost shield-like appearance.
Asaphid trilobites first appeared in the Late Cambrian (approximately 490 million years ago) and persisted into the Silurian Period, though they reached their greatest diversity and abundance during the Ordovician, before declining toward the end of that period around 444 million years ago.
One of the most striking features of Asaphida is the wide variation in eye development. Some genera possessed very large, well-developed eyes, suggesting an active lifestyle in well-lit, shallow marine environments. Others show reduced or completely absent eyes, an adaptation consistent with deeper-water or low-light habitats where vision was less critical.
Certain asaphids, most famously Isotelus, attained enormous sizes, making them among the largest trilobites ever to have lived. These giants were likely slow-moving benthic animals, possibly functioning as sediment sifters or detritivores, moving across the seafloor in search of organic material. Other members of the order display features that suggest adaptations to softer substrates or deeper marine settings.
The order Phacopida includes some of the most iconic and easily recognizable trilobites in the fossil record. Phacopid trilobites are best known for their distinctive schizochroal eyes, which are composed of large, individually separated calcite lenses, each with its own cornea. This highly advanced visual system likely provided excellent depth perception and strong sensitivity to changing light conditions, making phacopids among the most visually capable arthropods of the Paleozoic seas.
Phacopids first appeared during the Ordovician Period (approximately 485 million years ago) and persisted until the end of the Devonian Period, becoming extinct around 359 million years ago. They reached their greatest diversity and abundance during the Silurian and Devonian, when many of the best-known genera flourished.
These trilobites typically possessed robust, well-calcified bodies and were capable of tight, defensive enrollment, often forming nearly perfect spheres that protected their softer ventral surfaces. This ability, combined with their strong vision, suggests that phacopids were active benthic animals—likely scavengers or opportunistic predators—well adapted to life on the seafloor.

Pedinopariops, a Middle Devonian phacopid trilobite from Morocco, showcasing the classic robust form and prominent eyes typical of the order.
The order Lichida includes some of the most visually striking trilobites ever discovered. Lichid trilobites are characterized by heavily ornamented exoskeletons covered in prominent tubercles, nodes, and often spectacularly developed spines. In many species, these spines extend from the cephalon, thorax, and pygidium, giving lichids an unmistakably armored appearance.
Lichids first appeared in the Ordovician Period (approximately 485 million years ago) and reached their greatest diversity during the Silurian and Devonian, before disappearing by the end of the Devonian around 359 million years ago. These trilobites were likely benthic predators or scavengers, living on or near the seafloor where their robust bodies and spiny defenses would have offered protection from increasingly effective marine predators. Their dramatic morphology is widely interpreted as an evolutionary response to the escalating “arms race” of the Silurian–Devonian seas, a time when jawed fishes and other predators were becoming more abundant.

A classic, North American lichid trilobite (Arctinurus boltoni) from the Silurian deposits of New York.
The order Odontopleurida includes some of the most dramatically ornamented trilobites in the fossil record, best known for their extreme spinosity. Members of this order often possess long, slender, and delicate spines projecting from the cephalon, thorax, and pygidium, giving them a striking and highly defensive appearance.
Odontopleurid trilobites first appeared during the Late Ordovician Period (approximately 458 million years ago) and persisted until the Late Devonian, becoming extinct around 372 million years ago. They reached their greatest diversity and morphological elaboration during the Silurian and Devonian, when many of the most spectacular species evolved.
The exaggerated spines of odontopleurids are widely interpreted as adaptations against predation, making these trilobites difficult to grasp or consume by increasingly capable marine predators. In addition to defense, the spines may have served secondary functions, such as stabilization on soft or muddy substrates or increasing the animal’s apparent size.
Odontopleurid trilobites were benthic animals, living on or near the seafloor, and are generally thought to have been slow-moving scavengers or detritivores. Despite their elaborate defenses, they were unable to survive the ecological disruptions of the Late Devonian, when many heavily specialized trilobite groups declined or vanished.

The Odontopleurid trilobite (Kettneraspis williamsi) from the Haragan Formation of Oklahoma showing the long spines typical of the order.
The order Proetida represents the longest-surviving lineage of trilobites, persisting for more than 200 million years. Proetid trilobites first appeared in the Ordovician Period (approximately 485 million years ago) and survived until the end of the Permian Period, becoming extinct around 252 million years ago during the Permian–Triassic mass extinction. This event marked the final extinction of the entire trilobite class.
Members of Proetida are generally small to medium-sized trilobites with a conservative, streamlined body plan. They typically possess a well-defined cephalon, a thorax with a moderate number of segments, and a proportionally small pygidium. Compared to many earlier trilobite groups, proetids tend to show less extreme spinosity or ornamentation, a trait that may have contributed to their long-term evolutionary stability.
Proetid trilobites were ecological generalists, capable of inhabiting a wide range of marine environments. They are commonly interpreted as benthic detritivores or scavengers, living on or within seafloor sediments and feeding on organic material. Their flexibility in habitat and diet likely played a key role in their survival long after most other trilobite orders had vanished by the end of the Devonian.

A classic Proetid trilobite (Gerastos) from Morocco.
The group Agnostida was historically classified as a trilobite order and is still commonly referred to as such in older literature. However, most modern studies no longer recognize Agnostida as true trilobites.
Agnostids are extremely small, typically blind, and possess only two thoracic segments. Current research suggests they are more closely related to other arthropod groups, possibly within the stem lineage of crustaceans, rather than true members of Trilobita. As a result, Agnostida is generally excluded from modern trilobite classifications, though the topic remains an area of ongoing research.

Agnostids were historically classified as a trilobite order but are no longer considered to be true trilobites.
Although trilobites went extinct hundreds of millions of years before humans appeared, their fossils have been encountered by people for as long as humans have interacted with stone. Archaeological and historical evidence suggests that ancient cultures noticed, collected, and sometimes attributed meaning to trilobite fossils long before modern science understood what they were.
In parts of ancient North Africa and the Middle East, trilobite fossils may have been used as amulets or talismans, valued for their symmetry and unusual appearance. Some researchers have suggested that trilobites were incorporated into jewelry or carried as protective objects, though direct evidence is rare. In medieval Europe, trilobites were sometimes interpreted as petrified insects, snakes, or mythical creatures, and were occasionally called “stone beetles.”
In the Americas, certain Native American cultures are believed to have collected trilobite fossils, possibly as curiosities or spiritual objects. While oral traditions are difficult to tie conclusively to specific fossils, trilobites’ distinctive shapes would have made them stand out among naturally occurring stones.
In the modern world, trilobites occupy a unique space at the intersection of science, history, and art. What began centuries ago as curiosity-driven collecting has evolved into a global community of professional paleontologists, skilled fossil preparators, serious private collectors, and enthusiastic amateurs, all united by fascination with these ancient arthropods.
Today, trilobites are collected legally and responsibly from fossil-bearing rocks on every continent, with particularly important material coming from regions such as Morocco, the western United States, Russia, China, and parts of Europe. Fieldwork may involve carefully splitting shale layers by hand, extracting limestone blocks for later preparation, or conducting formal scientific excavations tied to research institutions.
For collectors, trilobites offer an unparalleled combination of diversity and individuality. No two specimens are exactly alike: each preserves a moment in deep time, whether a perfectly enrolled defensive posture, a molted exoskeleton left behind on the seafloor, or a fully articulated animal frozen in life position. Some collectors focus on classic localities and species, while others seek dramatic spiny forms, rare growth stages, or exceptionally prepared three-dimensional specimens.
Modern trilobite collecting also relies heavily on collaboration. Many scientifically important discoveries have been made by private collectors working closely with researchers, contributing specimens that expand knowledge of trilobite anatomy, development, and behavior. Museums and universities frequently acquire specimens from private collections, ensuring their long-term preservation and study.
While some trilobites are left in their natural, as-found state, others undergo meticulous preparation to reveal fine anatomical detail—transforming rough stone into striking windows onto Paleozoic life.
Over 20,000 described species of trilobites are known, and new species continue to be discovered. They ranged in size from tiny forms only a few millimeters long to giants exceeding 70 centimeters (over 2 feet), such as Isotelus rex, the largest trilobite ever found. Trilobites occupied nearly every marine ecological niche imaginable, from shallow coastal waters to deep-sea environments.

Trilobites: Ancient Arthropods of the Paleozoic Seas
Their fossils are so abundant and diverse that trilobites are often called the “index fossils of the Paleozoic.” Geologists use them to date and correlate rock layers across continents. Despite their long success and complexity, trilobites left no direct living descendants, making them a fully extinct branch of the arthropod tree. Beyond their scientific value, trilobites are prized by collectors for their intricate exoskeletons, dramatic spines, compound eyes, and exceptional preservation.
What makes trilobites especially fascinating is how much they reveal about early animal life: the evolution of complex eyes, the development of segmented bodies, and the rise of advanced behaviors hundreds of millions of years before vertebrates dominated the seas.
Scientists recognize 8 or 9 major trilobite orders, depending on classification scheme. Commonly recognized orders include Redlichiida, Ptychopariida, Asaphida, Phacopida, Lichida, Odontopleurida, Corynexochida, Harpetida, and Proetida, reflecting the extraordinary evolutionary diversity of the group.
What Is a Trilobite?
Trilobites are extinct members of the phylum Arthropoda, the same broad group that includes insects, spiders, crabs, and lobsters. They belong to their own class, Trilobita, and are not direct ancestors of any living animal, though they share many traits with modern arthropods. The name trilobite comes from the Greek tri (three) and lobos (lobe), referring to the three longitudinal sections of their bodies.

Trilobite Morphology
Trilobite anatomy is both highly standardized and remarkably adaptable, reflecting their long evolutionary history and ecological diversity. Their hardened exoskeleton was composed primarily of calcite and chitin, providing protection, structural support, and a surface for muscle attachment.
The Three Body Regions
Trilobite bodies are divided into three main regions:
Cephalon (Head)
The cephalon, or head, functioned as the control center of the trilobite. It typically housed a pair of compound eyes when present, a single pair of antennae used for sensory perception, and the mouth along with its associated feeding appendages. The central raised area of the cephalon is known as the glabella, which often overlies the stomach and digestive organs. Variations in the shape of the glabella are important for identifying trilobite species and may also reflect differences in feeding strategy.
Thorax (Articulated Body Segments)
The thorax consisted of multiple articulated segments, ranging from just a few to more than twenty depending on the species. These segments allowed flexibility for walking, burrowing, and enrolling. Each thoracic segment bore a pair of limbs beneath it.
Pygidium (Tail)
The pygidium was formed from fused segments and varied greatly in size. In some trilobites it was tiny (micropygous), while in others it was large or even larger than the head (macropygous). Its size and shape are key classification features.
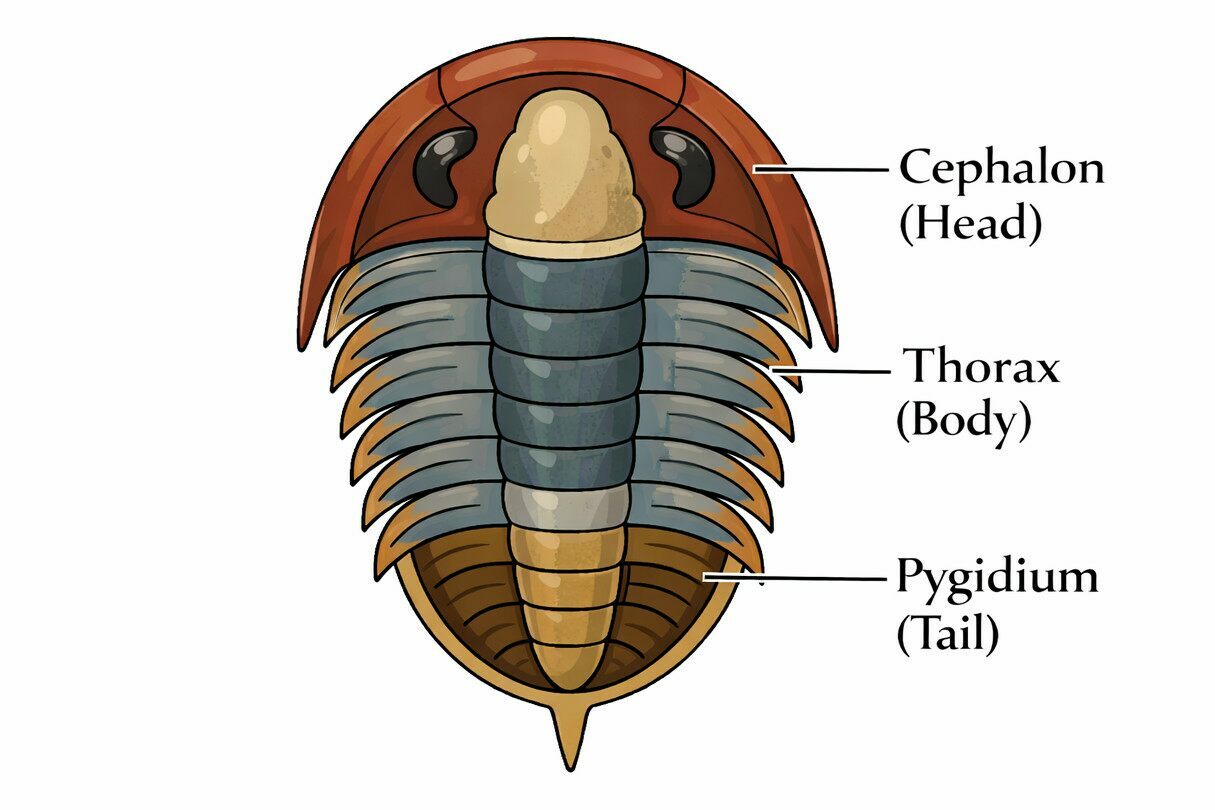
Trilobite Body Plan: Cephalon, Thorax and Pygidium
The Three Lobes
Viewed from above, trilobites are divided lengthwise into:
This tri-lobed arrangement is one of the defining features of the group.
Legs and Appendages
For much of scientific history, trilobite limbs were unknown, but exceptional fossil preservation has revealed that trilobites possessed biramous (two-branched) appendages, similar to those of some modern crustaceans.
Each body segment typically bore a pair of limbs consisting of:
These limbs suggest that trilobites were capable walkers on the seafloor and, in some cases, weak swimmers. Limb structure was surprisingly conservative across trilobite groups, even as exoskeleton shapes diversified.
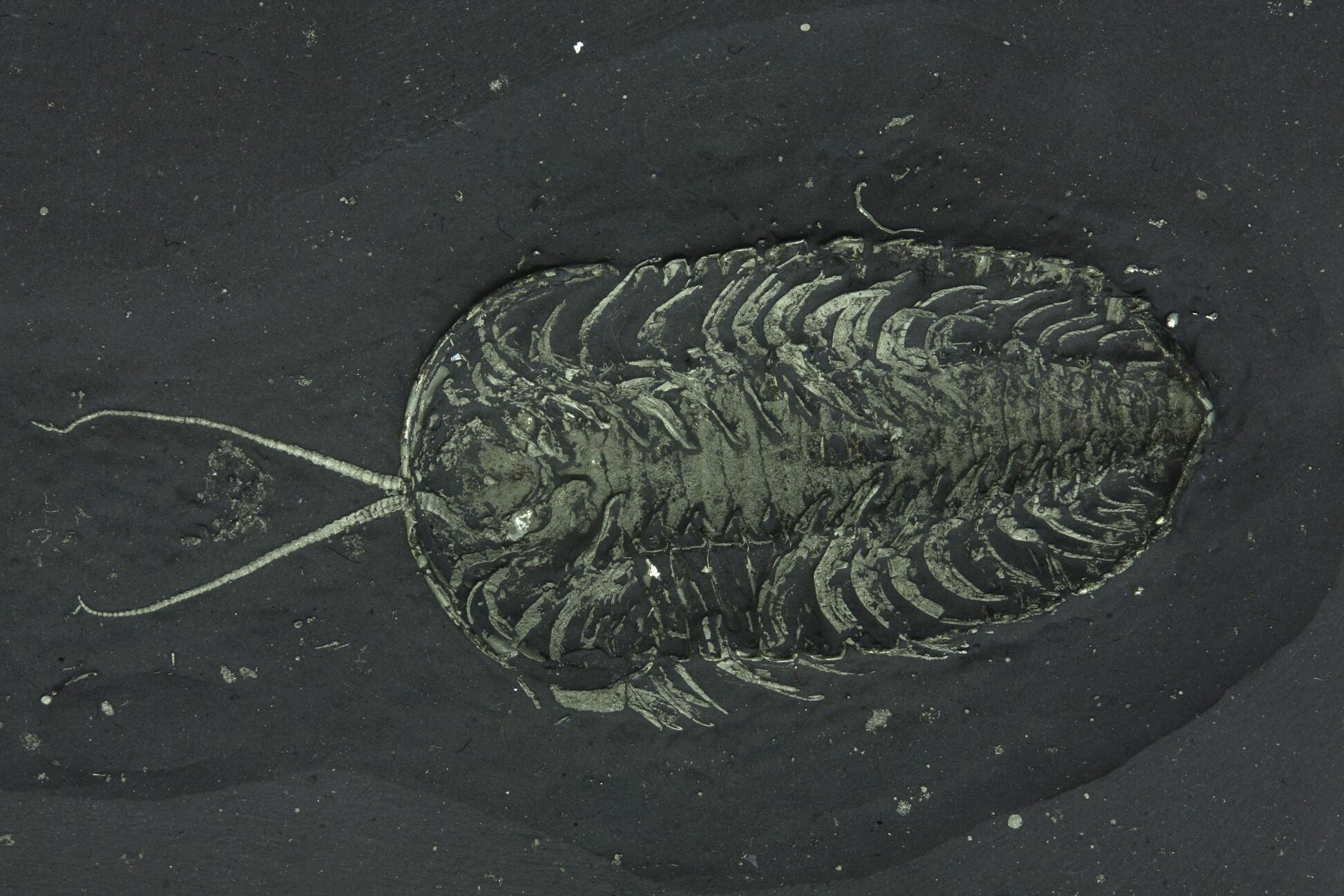
A ventrally preserved, Triarthrus trilobite from New York with rare soft-bodied preservation showing legs, gills and antennae.
Antennae and Sensory Structures
Trilobites had a single pair of antennae attached beneath the cephalon. These flexible, segmented structures were likely used to sense the environment, locate food, and navigate the seafloor. Antennae are rarely preserved but are known from sites such as the Burgess Shale and Chengjiang Biota.
Eyes: Among the Earliest Complex Eyes
Trilobite eyes are among the earliest complex eyes known in the fossil record. Most species possessed compound eyes made of calcite lenses—a unique feature rarely seen in modern animals. A few modern exceptions include brittlestars (ophiuroids), which use calcite lenses in their skeletal plates to sense light, and certain deep-sea crustaceans, which incorporate calcite into eye structures for light guidance rather than true image formation.
There were several eye types:
Some trilobites were completely blind, suggesting life in deep water or low-light environments.

A schizochroal eye of the trilobite Eldredgeops rana crassituberculata of Ohio.
Spines and Ornamentation
One of the most visually striking aspects of trilobite morphology is the presence of spines, tubercles, ridges, and other forms of ornamentation. These features vary enormously between groups and became increasingly elaborate over time.
Early Cambrian trilobites were generally smooth-bodied or only lightly ornamented. Prominent spines became more common during the Ordovician and especially widespread in the Silurian and Devonian, coinciding with the rise of increasingly effective predators such as jawed fishes and large cephalopods.
Possible Functions of Spines
Trilobite spines likely served multiple purposes, including:

A super spiny Comura bultyncki trilobite from Morocco covered in long spines.
Trilobites Known for Spines and Ornamentation
Some trilobite groups are especially famous for their elaborate armor:
In contrast, smooth-bodied trilobites such as many asaphids and proetids relied more on enrollment, size, or ecological positioning for protection rather than extreme ornamentation.
The evolution of spines and ornamentation in trilobites provides a clear example of an evolutionary "arms race" between predators and prey during the Paleozoic, recorded in stone across hundreds of millions of years.
Coloration and Appearance in Life
The original coloration of trilobites is not preserved in most fossils, as pigments break down during fossilization. However, paleontologists can make informed inferences about trilobite color based on comparisons with modern arthropods, ecological context, and rare exceptional preservation.
Most trilobites were likely colored in subdued tones such as browns, greens, grays, or mottled patterns that would have provided camouflage against the seafloor. Species living in shallow, well-lit environments may have exhibited more complex patterning to break up their outline, while deep-water or burrowing trilobites were likely uniformly dark or pale.
Some researchers have suggested that certain trilobites—particularly spiny or visually prominent forms—may have displayed contrasting patterns or highlights, either for species recognition or deterrence. However, bright warning coloration is considered unlikely for most species, given their benthic lifestyles and reliance on camouflage.
Rare fossil sites preserve subtle evidence of original surface texture, and in a few cases, differences in mineral replacement hint at possible pigment distribution, though these interpretations remain speculative.
Overall, trilobites were probably not brightly colored in the way many modern reef animals are, but instead displayed practical, environment-specific coloration well suited to survival in Paleozoic seas.
Key Anatomical Terms and Definitions
Growth and Molting
Like all arthropods, trilobites possessed a rigid, mineralized exoskeleton that could not grow continuously. To increase in size, they underwent a process known as molting (ecdysis), in which the old exoskeleton was periodically shed and replaced with a larger one. This occurred many times throughout a trilobite’s life, from tiny larval stages to full adulthood.
Molting in trilobites typically involved the opening or separation of specific parts of the exoskeleton—most commonly along facial sutures on the head or at the junction between the head (cephalon) and thorax. The trilobite would wriggle free of the old shell, leaving behind an empty exoskeleton that often retained its original shape. After emerging, the animal’s new exoskeleton was initially soft and vulnerable, hardening over time as it mineralized.
Many trilobite fossils actually represent molted exoskeletons rather than dead individuals, which helps explain their remarkable abundance in the fossil record. These molts are often recognizable by disarticulated parts, missing cheeks, or repeated occurrences of similarly sized specimens in the same layers.
Trilobites continued molting until reaching their final growth stage, after which they underwent a terminal or final molt. This last molt marked the end of growth and the onset of full maturity. In some species, the final molt coincided with changes in body proportions or the development of specialized features, such as exaggerated spines or ornamentation, possibly related to reproduction or defense. Once the final molt was complete, the trilobite would no longer shed its exoskeleton for the rest of its life.
How Paleontologists Identify Molts vs. Carcasses
Paleontologists distinguish trilobite molts from actual carcasses by examining how the fossil is preserved. Molted exoskeletons often show signs of deliberate separation, such as missing free cheeks, an opened head shield, or a gap between the head and thorax—common exit points used during molting. These remains may also appear flattened, slightly disarticulated, or repeated in large numbers at a single site, reflecting normal growth rather than mass death.
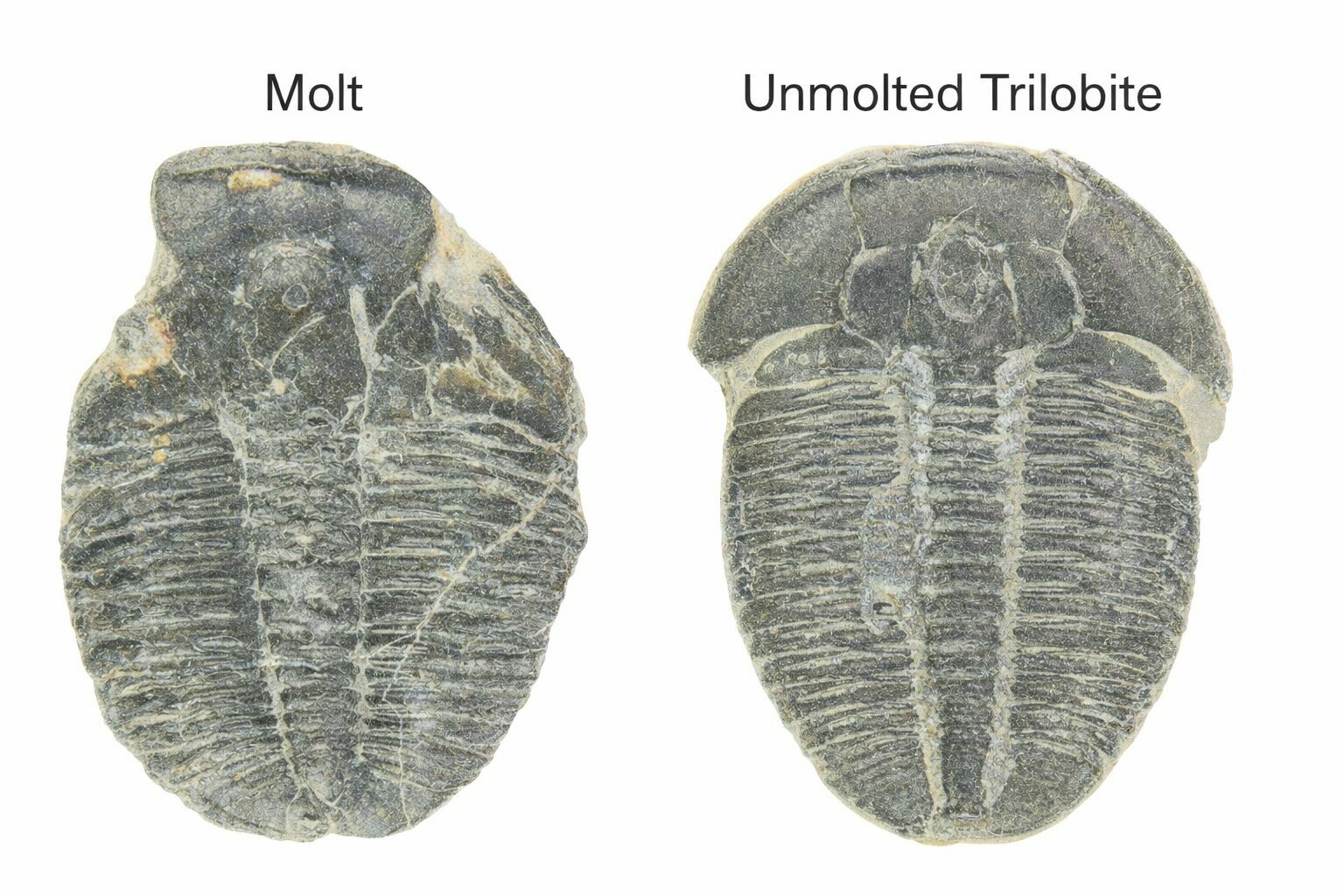
Two Elrathia trilobite fossils, the one on the left being a molt while the one on the right is the actual dead individual.
In contrast, trilobite carcasses are more likely to be fully articulated, with the head, thorax, and tail preserved in life position. Some show evidence of enrollment (curling up), a defensive posture unlikely during molting. The surrounding sediment and orientation can also provide clues, helping scientists determine whether a specimen represents a shed exoskeleton or the preserved remains of the animal itself.
Behavior and Ecology
Trilobites occupied a remarkable range of ecological roles on the ancient seafloor, with their modes of life closely intertwined with how they obtained food. Although the majority of trilobites were benthic—living on or just beneath the seafloor—their lifestyles were far from uniform, reflecting millions of years of adaptation to different marine environments.
Many trilobites lived as epifaunal crawlers, spending their lives moving across the surface of the sediment. These forms, common among groups such as ptychopariids and early redlichiids, likely foraged constantly as they crept over the seafloor. They fed primarily as scavengers and detritivores, consuming organic debris, decaying organisms, and microbial mats that blanketed Cambrian and Ordovician seabeds. Their anatomy was well suited to this lifestyle: moderately flexible bodies allowed them to navigate uneven substrates, while their limbs could manipulate sediment and food particles beneath them.
Other trilobites adopted a more concealed existence as shallow infaunal burrowers. These species were adapted to pushing through soft sediments, where they may have searched for buried organic material or small invertebrates. Smooth exoskeletons, reduced spines, and streamlined outlines helped minimize resistance as they moved beneath the surface. Living partially buried would have offered protection from predators while granting access to nutrient-rich sediments, making detritivory and opportunistic scavenging particularly effective feeding strategies in this niche.
A smaller but significant number of trilobites appear to have ventured above the seafloor. Some asaphids and other forms may have been capable of short bursts of swimming or hovering just above the substrate. Flattened bodies and well-developed pleural lobes would have aided this semi-nektonic lifestyle. These trilobites may have combined benthic feeding with the ability to relocate quickly or exploit suspended food resources, such as fine organic particles in the water column.

Cyclopyge is a rare, free-swimming trilobite distinguished by its enormous, downward-facing eyes. Its sleek, streamlined body indicates an active life in the open water column instead of along the ocean floor.
While scavenging and detritivory were the most widespread feeding strategies, trilobites were not limited to passive feeding. Certain robust and heavily ornamented species—particularly among the lichids—likely acted as active predators. With strong cephalic structures, spines for grasping or defense, and powerful appendages, these trilobites may have hunted small worms, soft-bodied arthropods, or other vulnerable invertebrates on or within the sediment.
Some trilobites may also have practiced filter feeding, using specialized appendages to strain suspended particles from the water, especially in calm, shallow marine environments. This strategy would have complemented a more stationary or slow-moving lifestyle, allowing trilobites to exploit nutrient-rich waters without constant locomotion.
Underlying these ecological roles was a strong link between form and function. The shape of the glabella and cephalon is thought to reflect feeding behavior, with broader or more expanded glabellas often associated with more active or specialized feeding strategies. Together, these anatomical and behavioral adaptations reveal trilobites not as uniform, primitive arthropods, but as highly diverse and ecologically sophisticated animals that played many roles in Paleozoic marine ecosystems.
Enrollment and Defensive Behavior
Enrollment—curling the body into a tight ball—was one of the most important defensive behaviors in trilobites. This behavior protected vulnerable soft tissues and concealed limbs beneath the exoskeleton. Trilobites with well-developed articulation and reinforced margins, such as phacopids, were especially adept at enrolling.
Many species also evolved long spines, thick armor, or ornamented exoskeletons, which served to deter predators or make handling more difficult.

A perfectly enrolled Morocops trilobite.
Predators of Trilobites
Trilobites were a major food source in Paleozoic marine ecosystems and faced predation throughout their history. Likely predators included:
Fossil evidence such as repaired injuries, bite marks, and abnormal growth patterns provides direct insight into predator–prey interactions.

A Utaspis trilboite from Utah with a large bite mark. Possibly left by an Anomalocaridid.
Social Behavior and Migration
Some fossil deposits contain mass mortality layers of trilobites, suggesting group behavior such as seasonal migration, synchronized molting, or breeding aggregations. Trackways preserved in sediment show multiple individuals moving in the same direction, supporting the idea of coordinated movement.
Such behaviors may have helped trilobites avoid predators, find mates, or exploit seasonal food resources.

A line of Lonchodomas trilobites showing potential migrating behavior.
Reproduction and Evidence from the Fossil Record
Direct evidence of trilobite reproduction is rare, but several important discoveries—particularly from Silurian and Ordovician deposits in New York State—provide valuable insight. Fossils from New York have yielded clusters of small, rounded structures preserved in close association with trilobite specimens, most notably with enrolled individuals of genera such as Flexicalymene. These clusters are widely interpreted as eggs, based on their size, shape, repeated occurrence, and spatial relationship to the adult trilobites.
In addition, exceptionally preserved specimens from New York have been interpreted to show paired internal structures consistent with reproductive organs, suggesting that some trilobites may have carried or brooded eggs rather than releasing them freely into the water column. This supports the idea that at least some trilobites practiced parental investment, a behavior seen in certain modern arthropods.
Together, these fossils indicate that trilobite reproduction was likely more complex than once assumed, involving egg production, possible brooding, and seasonal reproductive behavior tied to migration or molting cycles.
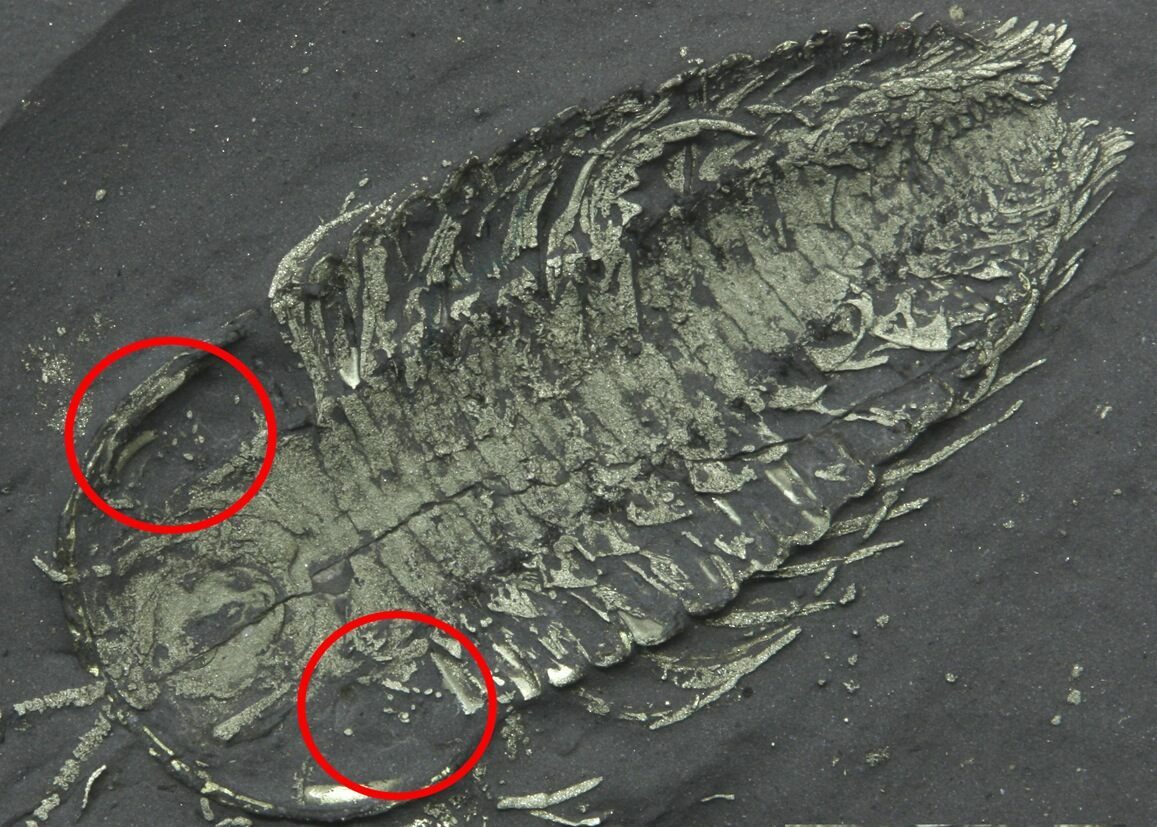
A triarthrus trilobite with what has been interpretted as eggs. Interestingly these are in the same location of the body where horseshoe crabs carry their eggs.
The Trilobite Fossil Record
Trilobites possess one of the richest and most informative fossil records of any extinct animal group. Their mineralized exoskeletons fossilized readily, and because they were abundant, widespread, and rapidly evolving, trilobites are invaluable for understanding Paleozoic geology and life. Trilobite fossils are found on every continent, including Antarctica, reflecting the global distribution of ancient seas.
Modes of Preservation: Flattened vs. Three-Dimensional
The way trilobites are preserved depends largely on sediment type, burial conditions, and post-depositional geology.
Exceptional preservation can also occur when trilobites are rapidly buried by sediment flows or preserved in low-oxygen environments that inhibit decay.
Important Trilobite Localities
Trilobite fossils have a truly global footprint, with discoveries spanning every continent and representing hundreds of millions of years of Earth’s history. From ancient seabeds preserved in fine detail to fossil-rich layers that capture extraordinary diversity, certain trilobite localities stand out above the rest. In this article, we highlight some of the world’s most important trilobite sites, chosen for the exceptional preservation of their specimens, the remarkable variety of species they yield, or their long-standing significance to fossil collectors and researchers alike.
Morocco
Morocco is widely regarded as the global epicenter of modern trilobite collecting and preparation, earning its reputation as the mecca for trilobite enthusiasts. The Anti-Atlas Mountains are especially famous for producing extraordinarily ornate Devonian trilobites, including large phacopids, lichids, and spiny odontopleurids that rank among the most visually striking trilobites ever discovered. In addition to these Devonian faunas, Morocco also hosts extensive Ordovician and Cambrian deposits, yielding large asaphids, early trilobite forms, and a remarkable continuity of trilobite history across much of the Paleozoic.

What makes Morocco truly unique is the combination of geology and human tradition. Vast regions of the Anti-Atlas expose fossil-bearing strata over enormous distances, with minimal vegetation, erosion cover, or tectonic faulting, allowing trilobite-rich layers to be traced and worked for miles. Thousands of people are involved in the commercial excavation and preparation of trilobites, often using highly refined techniques to extract fossils from hard limestone and reveal exquisite detail. As a result, Morocco is the source of the majority of trilobite specimens available on the global fossil market today, making it unmatched in both productivity and influence within the trilobite collecting community.
Burgess Shale - British Columbia, Canada
The Burgess Shale occupies a foundational place in the history of paleontology and trilobite research. Discovered in 1909 by Charles D. Walcott, this Cambrian fossil deposit—dated to approximately 508 million years ago—was among the first sites to demonstrate that delicate soft tissues could be preserved alongside mineralized skeletons. At the time of its discovery, this challenged long-held assumptions about the limits of fossil preservation and dramatically expanded scientific understanding of early animal life. Specimens preserving appendages, digestive systems, and respiratory structures provided the first compelling evidence that trilobites were complex, fully articulated animals rather than simple, rigid shells. These fossils laid the groundwork for modern interpretations of trilobite locomotion, feeding behavior, and ecological roles during the Cambrian explosion.
Today, the Burgess Shale is strictly protected, and collecting is prohibited, with material limited to museum collections and historically excavated specimens.
Great Basin, USA (Utah & Nevada)
The Great Basin region of Utah and Nevada is one of the most important trilobite areas in the world, with hundreds—if not thousands—of Cambrian species described, reflecting an extraordinary diversity during the early evolution of trilobites. Well-known Middle Cambrian deposits such as the Wheeler Shale and Marjum Formation in Utah’s House Range, along with Lower Cambrian strata near Pioche, Nevada, have been studied for over a century and continue to be actively collected by hundreds of amateur and professional fossil hunters each year.
Oklahoma, USA
Oklahoma holds a special place in trilobite collecting history, best known for its exquisitely preserved Devonian trilobites from the Haragan Formation, which have been collected and studied for decades. Localities such as Black Cat Mountain have produced spectacularly spiny odontopleurids and amazing phacopids, alongside a diverse assemblage of other Devonian forms, while lesser-known Ordovician and Silurian deposits reveal a surprisingly rich trilobite fauna spanning much of the Paleozoic. Oklahoma is also significant for its role in the evolution of fossil preparation, as Bob Carroll helped develop and popularize modern trilobite preparation techniques there—methods that have since become standard practice and elevated trilobite collecting into both a scientific and artistic discipline.

Collecting trilobites from the Haragan Formation in Oklahoma.
St. Petersburg Area, Russia
The Ordovician trilobite localities near St. Petersburg, Russia are among the most important classical trilobite sites in the world and have played a major role in the early development of trilobite research and taxonomy. Dating to roughly 485–444 million years ago, these deposits—often preserved in limestone and shale—have yielded abundant and well-studied trilobites, particularly large asaphid forms such as Asaphus, Megistaspis, and related genera. Many of these trilobites are notable for their broad, smooth bodies and, in some species, dramatically elevated eyes adapted for low-light environments.
These Russian Ordovician localities were heavily collected and described during the 19th and early 20th centuries, forming the basis for much of the foundational literature on Ordovician trilobites. Their exceptional abundance and preservation have made them critical for biostratigraphy and evolutionary studies, especially in understanding early trilobite gigantism and morphological specialization.
Upstate New York
New York State is one of the most historically important regions in the world for trilobite research, with classic fossil localities spanning the Ordovician and Silurian periods. The state is home to the famous Beecher’s Trilobite Beds (Upper Ordovician, ~445 million years old), a globally significant Lagerstätte where trilobites such as Triarthrus eatoni are preserved with soft tissues, including legs and antennae, through pyritization. These specimens provided some of the earliest direct evidence of trilobite limb anatomy and were critical to understanding trilobites as living arthropods rather than simple mineralized shells. New York is also renowned for the Walcott–Rust Quarry, famous for producing beautifully preserved 3D trilobites like Isotelus, Ceraurus and Flexicalymene
Ontario, Canada
Ontario is one of the most important regions in North America for Ordovician trilobites, preserving rich marine faunas dating to roughly 470–450 million years ago. Limestone formations across southern Ontario have yielded abundant trilobites such as Isotelus, Flexicalymene, Ceraurus, and Asaphus, including Isotelus rex, one of the largest trilobite species ever discovered and the official provincial fossil of Ontario. These deposits record shallow, warm Ordovician seas and have played a significant role in understanding trilobite diversity, morphology, and early Paleozoic marine ecosystems.
Historically, Ontario was a productive region for both research and collecting, with many classic museum specimens originating from active quarries. However, collecting has become increasingly difficult in recent decades due to quarry closures, restricted access, and stricter regulations. As a result, new material is far less common today, and Ontario trilobites are now valued primarily for their scientific and historical importance rather than abundance.
Famous Collectors, Researchers, and Their Importance
Trilobites have fascinated scientists, collectors, and the public for more than two centuries, playing a central role in the development of paleontology itself. Early researchers laid the foundation for our understanding of these ancient arthropods, none more influential than Charles Doolittle Walcott, whose discovery of the Burgess Shale in 1909 revealed exquisitely preserved Cambrian trilobites and other soft-bodied organisms. Before his famous discovery of the Burgess Shale in 1909, Walcott conducted some of his earliest and most important trilobite research at the Walcott–Rust Quarry near Trenton Falls, New York. Walcott’s work dramatically expanded scientific views of early animal life and helped establish trilobites as key indicators of early evolutionary complexity.

Modern day collection of trilobites from the hard limestones of the Walcott-Rust Quarry in New York
Throughout the 19th and 20th centuries, trilobites became a focus for pioneering paleontologists studying evolution, extinction, and ancient marine ecosystems. Their rapid evolutionary turnover and wide geographic distribution made them ideal subjects for biostratigraphy, allowing scientists to correlate rock layers across continents and reconstruct Earth’s deep-time history with remarkable precision.
In the modern era, the field has been equally shaped by the contributions of private collectors, preparators, and dedicated amateurs, particularly in regions such as Morocco, Utah, New York, and the American Midwest. Advances in preparation techniques—especially fine mechanical air abrasion—have revealed delicate features like compound eyes, spines, enrollment mechanisms, and even traces of soft anatomy. Many of the world’s finest trilobite specimens, including fully articulated and three-dimensionally preserved examples, come from the efforts of skilled private preparators working closely with researchers and museums.
This collaboration between collectors, commercial fossil preparators, academic institutions, and museums has led to the discovery of numerous new species and a deeper understanding of trilobite anatomy, behavior, and ecology. Today, trilobites remain indispensable to scientific research, helping illuminate patterns of evolution, mass extinction, and environmental change across hundreds of millions of years.
Beyond their scientific value, trilobites hold immense cultural and aesthetic significance. Their extraordinary diversity—ranging from smooth, streamlined forms to wildly spined and ornamented species—has made them one of the most sought-after fossils in the world. Equal parts scientific tool, historical artifact, and natural work of art, trilobites continue to inspire curiosity and admiration, securing their place as one of the most iconic fossils ever discovered.

An amazingly prepared Erbenochile trilobite from Morocco showing that these extrodinary fossil arthropods can also be natural works of art.
Evolution and Extinction
Origins and Possible Ancestors
Trilobites first appeared abruptly in the early Cambrian Period as part of the Cambrian Explosion, a time of rapid evolutionary experimentation when most major animal groups first emerged. Although no single direct ancestor has been definitively identified, trilobites are generally thought to have evolved from earlier arthropod-like stem groups.
Potential ancestors include soft-bodied Cambrian arthropods such as Spriggina, Parvancorina, and other members of the broader arthropod stem lineage, though these relationships remain debated. Fossils from sites like the Chengjiang Biota show transitional arthropods with segmented bodies, paired appendages, and developing cephalization—key traits that would later be fully expressed in trilobites.

A fossil of Spriggina floundensi, a possible ancestor of trilobites that lived during the Late Ediacaran, approximately 555 million years ago.
The sudden appearance of trilobites with complex eyes and mineralized exoskeletons suggests that their lineage had already undergone significant evolutionary development before entering the fossil record.
Cambrian Diversification
During the Cambrian, trilobites rapidly diversified into numerous forms and ecological roles. Early orders such as Redlichiida and Ptychopariida dominated Cambrian seas. These trilobites exhibited a wide range of body sizes, segment counts, and spinosity, reflecting intense evolutionary experimentation. Cambrian trilobites were primarily benthic crawlers and scavengers, but even at this early stage they occupied multiple niches. Frequent molting and rapid reproduction likely contributed to their success.
Ordovician Radiation
The Ordovician Period marked a second major diversification event for trilobites. Many Cambrian groups declined or disappeared, while more derived orders such as Asaphida rose to prominence. Trilobites became larger on average and expanded into deeper and more offshore environments. This period also saw increasing specialization, including eye reduction in deep-water forms and enhanced visual systems in shallow-water species. Trilobites were major components of increasingly complex marine ecosystems during the Ordovician.
Effects of the End-Ordovician Extinction
The end-Ordovician mass extinction, driven largely by global cooling and sea-level fluctuations, caused significant trilobite losses. Many families and genera vanished, particularly those adapted to shallow, warm-water environments. However, several lineages survived by occupying deeper or more flexible ecological niches.
Silurian and Devonian Adaptations
Following the Ordovician extinction, trilobites rebounded modestly during the Silurian. Orders such as Phacopida and Lichida diversified, often evolving robust bodies, improved enrollment mechanisms, and elaborate spines.
The Devonian Period brought increased predation pressure from rapidly evolving jawed fishes. In response, many trilobites evolved heavier armor, tighter enrollment, and defensive ornamentation. Despite these adaptations, overall trilobite diversity declined throughout the Devonian. The Late Devonian extinction events eliminated many remaining trilobite groups, leaving only a few resilient lineages.

During the Devonian trilobites increasingly evolved long spines, likely a defensive mechanism against predators like jawed fish.
Late Paleozoic Decline and the Proetida
By the Carboniferous Period, trilobites were reduced to a handful of families within the order Proetida. These trilobites were generally small, conservative in form, and often inhabited deeper or more restricted marine environments. Proetids survived multiple environmental fluctuations, including changes in sea level and oxygen availability, demonstrating remarkable resilience despite low diversity.
The Permian–Triassic Mass Extinction
The final extinction of trilobites occurred during the Permian–Triassic mass extinction, approximately 252 million years ago—the most severe extinction event in Earth’s history. Massive volcanic eruptions, ocean acidification, widespread anoxia, and extreme climate change devastated marine ecosystems. All remaining trilobites, including the last proetids, were wiped out during this event. Unlike earlier extinctions, there was no recovery or rebound for the group.
Evolutionary Legacy
Although trilobites left no direct descendants, their evolutionary legacy is profound. They provide critical insight into the early evolution of arthropods, the development of complex sensory systems, and the long-term dynamics of extinction and survival. Their rise and fall illustrate both the power and fragility of evolutionary success over geological time.
Trilobite Orders
Trilobites are traditionally divided into a set of major orders based on overall body plan, eye structure, thoracic segmentation, and evolutionary relationships. Depending on the classification scheme used, scientists recognize eight primary trilobite orders, with some debate over additional or historically used groupings.
Redlichiida
The order Redlichiida includes some of the earliest and most primitive trilobites known from the fossil record. Redlichiids first appeared in the Early Cambrian (approximately 521 million years ago) and persisted into the Middle Cambrian, disappearing by around 509 million years ago as more derived trilobite groups rose to prominence. They were an important component of early Cambrian marine ecosystems during the initial radiation of trilobites.
Redlichiid trilobites are typically characterized by large, crescent-shaped cephalons, numerous thoracic segments, and relatively small pygidia, giving them an elongated, flexible body plan. Many species possess long, sweeping genal spines extending from the rear of the cephalon, features that may have provided stability on soft substrates or offered some protection from early predators.
These trilobites are generally interpreted as active benthic crawlers, moving across the seafloor in search of food. Their morphology suggests a mobile lifestyle, and they are thought to have been among the earliest trilobites to explore a variety of shallow marine environments. Redlichiids represent some of the earliest evolutionary experiments in trilobite body plans, laying the groundwork for later, more specialized forms.

A Redlichiid trilobite (Olenellus nevadensis) from the Lower Cambrian of Nevada.
Ptychopariida
The order Ptychopariida represents one of the most diverse, abundant, and long-ranging groups of trilobites in the fossil record. Ptychopariids first appeared in the Early Cambrian (approximately 521 million years ago) and persisted into the Late Ordovician, with some lineages extending into the earliest Silurian. The group reached its greatest diversity and ecological importance during the Cambrian Period, when trilobites dominated marine ecosystems worldwide.
Members of this order typically possess moderate-sized cephalons, well-developed thoraxes composed of numerous segments, and a generally simple, conservative body plan. While often lacking the extreme spinosity or ornamentation seen in other trilobite orders, this morphological consistency proved highly successful and allowed ptychopariids to exploit a wide range of environments.
Ptychopariid trilobites occupied diverse ecological niches, ranging from shallow continental shelf settings to deeper offshore environments. Most are interpreted as benthic detritivores or scavengers, living on or within soft seafloor sediments. Their adaptability and wide geographic distribution made them one of the most successful trilobite groups of the Paleozoic.
Because of their abundance, rapid evolutionary turnover, and broad distribution, ptychopariids are extremely important for biostratigraphy, particularly in Cambrian-aged rocks where they are widely used to define and correlate stratigraphic zones.

The Ptychopariid trilobite (Elrathia kingii) from the Wheeler Shale in Utah is probably the most recognizable trilobite in the world due to millions of them having been collected.
Corynexochida
The order Corynexochida includes a diverse assemblage of trilobites distinguished by their forward-tapering (conical) glabellas, a feature that often gives the cephalon a pointed or wedge-shaped appearance. Many corynexochids also bear prominent thoracic or cephalic spines, though the degree of ornamentation varies widely across the group.
Corynexochid trilobites first appeared during the Early Cambrian (approximately 521 million years ago) and persisted into the Ordovician Period, with some lineages extending into the early Silurian. The group reached its greatest diversity during the Cambrian, when trilobites were undergoing rapid evolutionary radiation, and remained an important component of marine faunas into the Ordovician before declining.
Morphologically, Corynexochida encompasses both smooth-bodied, streamlined forms and heavily ornamented species with long, sweeping spines, reflecting a wide range of ecological strategies. These trilobites are generally interpreted as active benthic animals, living on or near the seafloor. Some species show features consistent with predatory or scavenging behavior, while others may have been semi-infaunal, partially burrowing into soft sediments for feeding or protection.

A Corynexochid trilboite (Olenoides nevadensis) from the Middle Cambrian deposits of Utah.
Asaphida
The order Asaphida includes a diverse group of often large, smooth-bodied trilobites that were especially successful during the Ordovician Period. Asaphids are typically characterized by a wide, flattened body, a broad pygidium, and a reduced number of thoracic segments, giving many species a streamlined, almost shield-like appearance.
Asaphid trilobites first appeared in the Late Cambrian (approximately 490 million years ago) and persisted into the Silurian Period, though they reached their greatest diversity and abundance during the Ordovician, before declining toward the end of that period around 444 million years ago.
One of the most striking features of Asaphida is the wide variation in eye development. Some genera possessed very large, well-developed eyes, suggesting an active lifestyle in well-lit, shallow marine environments. Others show reduced or completely absent eyes, an adaptation consistent with deeper-water or low-light habitats where vision was less critical.
Certain asaphids, most famously Isotelus, attained enormous sizes, making them among the largest trilobites ever to have lived. These giants were likely slow-moving benthic animals, possibly functioning as sediment sifters or detritivores, moving across the seafloor in search of organic material. Other members of the order display features that suggest adaptations to softer substrates or deeper marine settings.

Phacopida
The order Phacopida includes some of the most iconic and easily recognizable trilobites in the fossil record. Phacopid trilobites are best known for their distinctive schizochroal eyes, which are composed of large, individually separated calcite lenses, each with its own cornea. This highly advanced visual system likely provided excellent depth perception and strong sensitivity to changing light conditions, making phacopids among the most visually capable arthropods of the Paleozoic seas.
Phacopids first appeared during the Ordovician Period (approximately 485 million years ago) and persisted until the end of the Devonian Period, becoming extinct around 359 million years ago. They reached their greatest diversity and abundance during the Silurian and Devonian, when many of the best-known genera flourished.
These trilobites typically possessed robust, well-calcified bodies and were capable of tight, defensive enrollment, often forming nearly perfect spheres that protected their softer ventral surfaces. This ability, combined with their strong vision, suggests that phacopids were active benthic animals—likely scavengers or opportunistic predators—well adapted to life on the seafloor.

Pedinopariops, a Middle Devonian phacopid trilobite from Morocco, showcasing the classic robust form and prominent eyes typical of the order.
Lichida
The order Lichida includes some of the most visually striking trilobites ever discovered. Lichid trilobites are characterized by heavily ornamented exoskeletons covered in prominent tubercles, nodes, and often spectacularly developed spines. In many species, these spines extend from the cephalon, thorax, and pygidium, giving lichids an unmistakably armored appearance.
Lichids first appeared in the Ordovician Period (approximately 485 million years ago) and reached their greatest diversity during the Silurian and Devonian, before disappearing by the end of the Devonian around 359 million years ago. These trilobites were likely benthic predators or scavengers, living on or near the seafloor where their robust bodies and spiny defenses would have offered protection from increasingly effective marine predators. Their dramatic morphology is widely interpreted as an evolutionary response to the escalating “arms race” of the Silurian–Devonian seas, a time when jawed fishes and other predators were becoming more abundant.

A classic, North American lichid trilobite (Arctinurus boltoni) from the Silurian deposits of New York.
Odontopleurida
The order Odontopleurida includes some of the most dramatically ornamented trilobites in the fossil record, best known for their extreme spinosity. Members of this order often possess long, slender, and delicate spines projecting from the cephalon, thorax, and pygidium, giving them a striking and highly defensive appearance.
Odontopleurid trilobites first appeared during the Late Ordovician Period (approximately 458 million years ago) and persisted until the Late Devonian, becoming extinct around 372 million years ago. They reached their greatest diversity and morphological elaboration during the Silurian and Devonian, when many of the most spectacular species evolved.
The exaggerated spines of odontopleurids are widely interpreted as adaptations against predation, making these trilobites difficult to grasp or consume by increasingly capable marine predators. In addition to defense, the spines may have served secondary functions, such as stabilization on soft or muddy substrates or increasing the animal’s apparent size.
Odontopleurid trilobites were benthic animals, living on or near the seafloor, and are generally thought to have been slow-moving scavengers or detritivores. Despite their elaborate defenses, they were unable to survive the ecological disruptions of the Late Devonian, when many heavily specialized trilobite groups declined or vanished.

The Odontopleurid trilobite (Kettneraspis williamsi) from the Haragan Formation of Oklahoma showing the long spines typical of the order.
Proetida
The order Proetida represents the longest-surviving lineage of trilobites, persisting for more than 200 million years. Proetid trilobites first appeared in the Ordovician Period (approximately 485 million years ago) and survived until the end of the Permian Period, becoming extinct around 252 million years ago during the Permian–Triassic mass extinction. This event marked the final extinction of the entire trilobite class.
Members of Proetida are generally small to medium-sized trilobites with a conservative, streamlined body plan. They typically possess a well-defined cephalon, a thorax with a moderate number of segments, and a proportionally small pygidium. Compared to many earlier trilobite groups, proetids tend to show less extreme spinosity or ornamentation, a trait that may have contributed to their long-term evolutionary stability.
Proetid trilobites were ecological generalists, capable of inhabiting a wide range of marine environments. They are commonly interpreted as benthic detritivores or scavengers, living on or within seafloor sediments and feeding on organic material. Their flexibility in habitat and diet likely played a key role in their survival long after most other trilobite orders had vanished by the end of the Devonian.

A classic Proetid trilobite (Gerastos) from Morocco.
Note on Agnostida
The group Agnostida was historically classified as a trilobite order and is still commonly referred to as such in older literature. However, most modern studies no longer recognize Agnostida as true trilobites.
Agnostids are extremely small, typically blind, and possess only two thoracic segments. Current research suggests they are more closely related to other arthropod groups, possibly within the stem lineage of crustaceans, rather than true members of Trilobita. As a result, Agnostida is generally excluded from modern trilobite classifications, though the topic remains an area of ongoing research.

Agnostids were historically classified as a trilobite order but are no longer considered to be true trilobites.
Trilobites and Humans
Although trilobites went extinct hundreds of millions of years before humans appeared, their fossils have been encountered by people for as long as humans have interacted with stone. Archaeological and historical evidence suggests that ancient cultures noticed, collected, and sometimes attributed meaning to trilobite fossils long before modern science understood what they were.
In parts of ancient North Africa and the Middle East, trilobite fossils may have been used as amulets or talismans, valued for their symmetry and unusual appearance. Some researchers have suggested that trilobites were incorporated into jewelry or carried as protective objects, though direct evidence is rare. In medieval Europe, trilobites were sometimes interpreted as petrified insects, snakes, or mythical creatures, and were occasionally called “stone beetles.”
In the Americas, certain Native American cultures are believed to have collected trilobite fossils, possibly as curiosities or spiritual objects. While oral traditions are difficult to tie conclusively to specific fossils, trilobites’ distinctive shapes would have made them stand out among naturally occurring stones.

Modern Trilobite Collecting
In the modern world, trilobites occupy a unique space at the intersection of science, history, and art. What began centuries ago as curiosity-driven collecting has evolved into a global community of professional paleontologists, skilled fossil preparators, serious private collectors, and enthusiastic amateurs, all united by fascination with these ancient arthropods.
Today, trilobites are collected legally and responsibly from fossil-bearing rocks on every continent, with particularly important material coming from regions such as Morocco, the western United States, Russia, China, and parts of Europe. Fieldwork may involve carefully splitting shale layers by hand, extracting limestone blocks for later preparation, or conducting formal scientific excavations tied to research institutions.
For collectors, trilobites offer an unparalleled combination of diversity and individuality. No two specimens are exactly alike: each preserves a moment in deep time, whether a perfectly enrolled defensive posture, a molted exoskeleton left behind on the seafloor, or a fully articulated animal frozen in life position. Some collectors focus on classic localities and species, while others seek dramatic spiny forms, rare growth stages, or exceptionally prepared three-dimensional specimens.
Modern trilobite collecting also relies heavily on collaboration. Many scientifically important discoveries have been made by private collectors working closely with researchers, contributing specimens that expand knowledge of trilobite anatomy, development, and behavior. Museums and universities frequently acquire specimens from private collections, ensuring their long-term preservation and study.
While some trilobites are left in their natural, as-found state, others undergo meticulous preparation to reveal fine anatomical detail—transforming rough stone into striking windows onto Paleozoic life.





